 |
 |
|
|
![]()
 Краткие
сведения о классификации растений
Краткие
сведения о классификации растений
Растения
ниших природных водоёмов

![]()
Происхождение растений
Растения как типичные представители фотоавтотрофных организмов нашей планеты возникли в ходе длительной эволюции, которая ведет свое начало от примитивных обитателей освещенной зоны моря - планктонных и бентосньгх прокариот. Сопоставляя палеонтологические данные с данными сравнительной морфологии и физиологии, ныне живущих растений, можно в общем, виде наметить следующую хронологическую последовательность их появления и развития:
-
бактерии и сине-зеленые водоросли (прокариоты);
-
водоросли циановые, зеленые, бурые, красные и др. (эукариоты, как и все последующие организмы);
-
мхи и печеночники;
-
папоротники, хвощи, плауны, семенные папоротники;
-
голосеменные (включая шишковые растения);
-
покрытосеменные, или цветковые, растения.
Бактерии и сине-зеленые водоросли обнаружены в наиболее древних сохранившихся отложениях докембрия, значительно позже появляются водоросли, и только в фанерозое мы встречаем пышное развитие высших растений: плауновых, хвощей, голосеменных и покрытосеменных.
В течение всего криптозоя в первичных водоемах в эвфотической зоне древних морей развивались преимущественно одноклеточные организмы - водоросли различного типа.
У основных представителей прокариот, обнаруженных в докембрии, питание было автотрофным - с помощью фотосинтеза. Наиболее благоприятные условия для фотосинтеза создавались в освещенной части моря па глубине от поверхности до 10 м, что соответствовало также условиям мелководного бентоса.
К настоящему времени изучение докембрийских микрофоссилий продвинулось вперед, соответственно накоплен большой фактический материал. В целом интерпретация микроскопических препаратов представляет собой трудную задачу, которую нельзя разрешить однозначно.
Лучше всего выявляются и опознаются трихомные бактерии, резко отличающиеся от минеральных образований близкой формы. Полученный эмпирический материал по микрофоссилиям позволяет заключить, что их можно сопоставить с ныне живущими цианобактериями.
Строматолиты, как биогенные постройки далекого прошлого планеты, образовались при накоплении тонкого осадка карбоната кальция, захватываемого фотосинтезирующими организмами микробиологических ассоциаций. Микрофоссилии в строматолитах состоят почти исключительно из прокариотических микроорганизмов, относящихся главным образом к сине-зеленым водорослям - цианофиты. При изучении остатков бентосных микроорганизмов, слагающих строматопиты, выяснилась одна интересная особенность, имеющая принципиальное значение. Микрофоссилии разного возраста мало изменяют свою морфологию и свидетельствуют в целом о консервативности прокариот. Микрофоссилии, относящиеся к прокариотам, оставались практически постоянными довольно долгое время. Во всяком случае, перед нами установленный факт - эволюция прокариот проходила значительно медленнее, чем высших организмов.
Итак, в ходе геологической истории бактерии-прокариоты обнаруживают максимальную персистентность. К персистентным формам относятся организмы, сохранившиеся в процессе эволюции в неизменном виде. Как отмечает Г. А. Заварзин [1984], поскольку древние сообщества микроорганизмов обнаруживают значительные черты сходства с современными, развивающимися в гидротермах и в областях образования эвапоритов, то это позволяет на современных природных илах. Итак, в ходе геологической истории бактерии-прокариоты обнаруживают максимальную персистентность. К персистентным формам относятся организмы, сохранившиеся в процессе эволюции в неизменном виде. Как отмечает Г. А. Заварзин [1984], поскольку древние сообщества микроорганизмов обнаруживают значительные черты сходства с современными, развивающимися в гидротермах и в областях образования эвапоритов, то это позволяет на современных природных и лабораторных моделях более обстоятельно изучить геохимическую деятельность этих сообществ, экстраполируя их в далекое докембрийское время.
Первые эукариоты возникли в планктонных ассоциациях открытых вод. Конец исключительного господства прокариот относится приблизительно к дате 1,4 млрд лет назад, хотя первые эукариоты появились значительно раньше. Так, по последним данным, облик ископаемых органических остатков из черных сланцев и углистых образований района Верхнего Озера свидетельствует о появлении эукариотических микроорганизмов 1,9 млрд лет назад.
От даты 1,4 млрд лет назад к нашему времени палеонтологическая летопись докембрия значительно расширяется. К этой дате приурочено появление сравнительно крупных форм, относящихся к планктонным эукариотам и получивших название ''акритархи'' (в переводе с греческого - ''существа неизвестного происхождения'' ). Следует отметить, что группа акритарх (Acritarcha) предложена в качестве неопределенной систематической категории, обозначающей Микрофоссилии различного происхождения, но сходные по внешним морфологическим признакам. В литературе описаны акритархи из докембрия и нижнего палеозоя. Большинство акритарх были, вероятно, одноклеточными фотосинтезирующими эукариотами - оболочками каких-то древних водорослей. Некоторые из них могли еще иметь прокариотическую организацию. На планктонный характер акритарх указывает их космополитическое распространение в одновозрастных осадочных отложениях. Наиболее древние акритархи из отложений раннего рифея Южного Урала обнаружил Т. В. Янкаускас.
В ходе геологического времени размеры акритарх увеличивались. По данным наблюдений, оказалось, что, чем моложе докембрийские Микрофоссилии, тем они крупнее. Допускается, что значительное увеличение размеров акритарх было связано с увеличением размеров эукариотической организации клеток. Они могли появиться как самостоятельные организмы либо, что более вероятно, в симбиозе с другими. Л. Маргелис считает, что эукариотические клетки скомпоновались из уже существовавших прокариотических. Однако для выживания эукариот необходимо было, чтобы среда обитания была насыщена кислородом и, как следствие этого, возник аэробный метаболизм. Первоначально свободный кислород, выделяющийся при фотосинтезе цианофитов, накапливался в ограниченных количествах в мелководных местах обитания. Возрастание его содержания в биосфере вызвало реакцию со стороны организмов: они стали заселять бескислородные места обитания (в частности, анаэробные формы).
Данные докембрийской микропалеонтологии указывают, что в среднем докембрии еще до появления эукариот цианофиты составляли относительно небольшую часть планктона. Эукариоты нуждались в свободном кислороде, и все более конкурировали с прокариотами в тех областях биосферы, где появлялся свободный кислород. По имеющимся данным микропалеонтологии можно судить, что переход от прокариотной к эукариотной флоре древних морей совершался медленно и обе группы организмов долгое время сосуществовали вместе. Впрочем, это сосуществование в иной пропорции происходит и в современную эпоху. К началу позднего рифея уже распространилось множество автотрофных и гетеротрофных форм организмов.
По мере своего развития организмы перемещались за питательными веществами в более глубокие и удаленные от шельфов области моря. В палеонтологической летописи отмечается резкое увеличение разнообразия крупных сфероидальных форм эукариотических акритарх в позднерифейское время, 900-700 млн лет назад. Около 800 млн лет назад в Мировом океане появились представители нового класса планктонных организмов - кубкообразные тельца с массивными раковинами или наружными покровами, минерализованными карбонатом кальция или кремнеземом. В начале кембрийского периода в эволюции планктона произошли существенные сдвиги - возникли разнообразные микроорганизмы со сложной скульптурной поверхностью и улучшенной плавучестью. Они дали начало настоящим шиповатьш акритархам.
Появление эукариот создало важную предпосылку для зарождения в раннем рифее (около 1,3 млрд лет назад) многоклеточных растений и животных. Для белтской серии из докембрия западных штатов Северной Америки их описал еще Ч. Уолкотт, Но к какому типу водорослей они относятся (бурые, зеленые или красные), еще неясно. Таким образом, чрезвычайно длительная эра господства бактерий и близких к ним сине-зеленых водорослей сменилась эрой водорослей, достигавших в водах древних океанов значительного разнообразия форм и красок. На протяжении позднего рифея и венда многоклеточные водоросли становятся разнообразнее, они сопоставляются с бурыми и красными.
По мнению академика Б. С. Соколова [1979], многоклеточные растения и животные появились почти одновременно. В отложениях венда встречаются разнообразные представители водных растений. Наиболее видное место занимают многоклеточные водоросли, слоевища которых нередко переполняют толщи вендских отложений: аргиллитов, глин, песчаников. Часто встречаются макропланктонные водоросли, колониальные, спиральные нитчатые водоросли Volymella, войлоковидные и другие формы. Весьма разнообразен фитопланктон.
В течение большей части истории Земли эволюция растений проходила в водной среде. Именно здесь зародилась и прошла различные этапы развития водная растительность. В целом водоросли - это обширная группа низших водных растений, содержащих хлорофилл и вырабатывающих органические вещества путем фотосинтеза. Тело водорослей еще не дифференцировано на корни, листья и другие характерные части. Они представлены одноклеточными, многоклеточными и колониальными формами. Размножение бесполое, вегетативное и половое. Водоросли входят в состав планктона и бентоса. В настоящее время их относят к подцарству растений Thallophyta, y которых тело сложено относительно однородной тканью - слоевищем, или Thallus. Слоевище состоит из многих клеток, сходных по своему виду и функциям. В историческом аспекте водоросли прошли наиболее длительный этап в развитии зеленых растений и в общем, геохимическом круговороте вещества биосферы сыграли роль гигантского генератора свободного кислорода. Возникновение и развитие водорослей носило крайне неравномерный характер.
Зеленые водоросли (Chlorophyta) представляют собой обширную и широко распространенную группу преимущественно зеленых растений, которая распадается на пять классов. По внешнему виду они сильно отличаются друг от друга. Зеленые водоросли происходят от зеленых жгутиковых организмов. Об этом свидетельствуют переходные формы - пирамидомонас и хламидомонас, подвижные одноклеточные организмы, обитающие в водах. Размножаются зеленые водоросли половым путем. Некоторые группы зеленых водорослей достигли большого развития в триасовый период.
Жгутиковые (Flagellata) объединяются в группу микроскопических одноклеточных организмов. Одни исследователи относят их к царству растений, другие - к царству животных. Подобно растениям, некоторые жгутиковые содержат хлорофилл. Однако в отличие от большинства растений они не имеют обособленной клеточной системы и способны переваривать пищу с помощью ферментов, а также жить в темноте, как животные организмы. По всей вероятности, жгутиковые существовали в докембрии, но бесспорные их представители найдены в юрских отложениях.
Бурые водоросли (Phaeophyta) отличаются присутствием бурого пигмента в таком количестве, что он фактически маскирует хлорофилл и придает растениям соответствующую окраску. Бурые водоросли относятся к бентосу и планктону. Самые крупные водоросли достигают 30 м в длину. Почти все они произрастают в соленой воде, поэтому их называют морской травой. К бурым относят саргассовые водоросли - плавающие планктонные формы с большим количеством пузырьков. В ископаемом состоянии известны с силура.
Красные водоросли (Rhodophyta) имеют такую окраску благодаря красному пигменту. Это преимущественно морские растения, сильно разветвленные. У некоторых из них известковый скелет. Эту группу часто называют куллипорами. Они существуют и в настоящее время, а в ископаемом состоянии известны с нижнего мела. Близкие к ним сомипоры, с более крупными и широкими клетками, появились в ордовике.
Харовые водоросли (Charophyta) представляют собой весьма своеобразную и довольно высокоорганизованную группу многоклеточных растений, размножающихся половым путем. Они настолько отличаются от других водорослей, что некоторые ботаники относят их к листостебельным ввиду наметившейся дифференциации тканей. Харовые водоросли зеленого цвета, обитают в настоящее время в пресной воде и в солоноватых водоемах. Они избегают морской воды с нормальной соленостью, но можно предположить, что в палеозое они населяли моря. У отдельных харофитов возникают споропочки, пропитанные углекислым кальцием. Харовые водоросли принадлежат к важным породообразующим организмам пресноводных известняков.
Диатомовые водоросли (Diatomeae) - типичные представители планктона. У них продолговатая форма, снаружи покрыты панцирем, состоящим из кремнезема. Первые остатки диатомовых водорослей найдены в отложениях девона, но возможно, они древнее. В целом диатомовые водоросли относительно молодая группа. Эволюция их изучена лучше, чем других водорослей, поскольку кремневые панцири и створки диатомей могут сохраняться в ископаемом состоянии очень длительное время. По всей вероятности, диатомовые водоросли происходят от жгутиковых, окрашенных в желтый цвет и способных отлагать в своих оболочках небольшое количество кремнезема. В современную эпоху диатомей широко распространены в пресных и морских водах, изредка встречаются и во влажных почвах. Остатки диатомей известны в юрских отложениях, однако, не исключено, что они появились значительно раньше. Ископаемые диатомовые водоросли от раннего мела дошли до современной эпохи без перерыва в отложениях.
Очень важным событием, способствовавшим резкому ускорению темпов эволюции всего живого населения нашей планеты, был выход растений из морской среды на сушу. Выход растений на поверхность континентов можно считать подлинной революцией в истории биосферы. Развитие наземной растительности создало предпосылку для выхода на сушу животных. Однако массовому переходу растений на сушу предшествовал длительный подготовительный период. Можно предполагать, что растительная жизнь на суше появилась очень давно, во всяком случае, локально - во влажном климате на побережьях мелководных заливов и лагун, где при изменениях уровня воды периодически происходило поступление водной растительности на сушу. Советский натуралист Л. С. Берг впервые высказал мысль, что поверхность суши не представляла собой безжизненной пустыни ни в кембрии, ни в докембрии. Видный советский палеонтолог Л. Ш. Давиташвили также допускал, что в докембрии на материках, вероятно, уже было какое-то население, состоящее из низкоорганизованных растений и, возможно, даже животных. Однако их общая биомасса была ничтожной.
Чтобы жить на суше, растения должны были не терять воду. Следует при этом иметь в виду, что у высших растений - мхов, папоротникообразных, голосеменных и цветковых, составляющих в настоящее время главную массу земной растительности, соприкасаются с водой только корни, корневые волоски и ризоиды, остальные же их органы находятся в атмосфере и испаряют воду всей поверхностью.
Наиболее расцвела растительная жизнь на берегах лагунных озер и болот. Здесь появился тип растения, нижняя часть которого находилась в воде, а верхняя - в воздушной среде, под прямыми лучами солнца. Несколько позже, с проникновением растений на незатопляемую сушу, самые первые их представители развили корневую систему и получили возможность потреблять грунтовые воды. Это способствовало их выживанию в засушливые периоды. Таким образом, новые обстоятельства привели к расчленению клеток растений на ткани и выработке защитных приспособлений, каких не было у обитавших в воде предков.

Рис.14. Развитие и генетические связи различных групп наземных растений
Массовое завоевание континентов растениями произошло в силурийский период палеозойской эры. Прежде всего, это были псилофиты - своеобразные споровые растения, напоминающие плауны. Часть извилистых стеблей псилофитов была покрыта щетинистыми листьями. Псилофиты были лишены корней, а в основном и листьев. Они состояли из ветвившихся зеленых стеблей высотой до 23 см и горизонтально тянувшегося в почве корневища. Псилофиты, как первые достоверные растения суши, создавали целые зеленые ковры на влажной почве.
Вероятно, продукция органического вещества первых растительных покровов суши была незначительной. Растительность силурийского периода, несомненно, произошла от водорослей моря, и сама породила растительность последующего периода.
После завоевания суши развитие растительности привело к образованию многочисленных и разнообразных форм. Интенсивное разделение растительных групп началось в девоне и продолжалось в последующее геологическое время. Общая родословная важнейших групп растений дана на рис. 14.
Мхи произошли от. водорослей. Их ранняя стадия развития очень сходна с некоторыми зелеными водорослями. Однако существует предположение, что мхи произошли от более простых представителей бурых водорослей, приспособившихся к жизни на сырых скалах или вообще в почвах.
На поверхности раннепалеозойских континентов век водорослей сменился веком псилофитов, давших растительность, напоминавшую по своему внешнему виду и размерам современные заросли крупных мхов. Господство псилофитов сменилось в каменноугольный период господством папоротникообразных растений, образовавших довольно обширные леса на болотистых почвах. Развитие этих растений способствовало тому, что состав атмосферного воздуха изменился. Добавилось существенное количество свободного кислорода, и накопилась масса пищевых веществ, необходимых для возникновения и развития сухопутных позвоночных животных. В то же время были накоплены огромные массы каменного угля. Каменноугольный период характеризовался исключительным расцветом наземной растительности. Возникли древовидные плауны, достигавшие в высоту 30 м, огромные хвощи, папоротники начали появляться хвойные. В пермский период продолжалось развитие наземной растительности, которая значительно расширила места своего обитания.
Период господства папоротникообразных сменился периодом шишконосных хвойных растений. Поверхность материков стала приобретать современный облик. В начале мезозойской эры большое распространение получили хвойные, цикадовые, а в меловой период появляются цветковые растения. В самом начале раннемеловой эпохи еще существовали юрские формы растений, но затем состав растительности сильно изменился. В конце раннемеловой эпохи встречается много покрытосеменных растений. С самого же начала позднемеловой эпохи они оттесняют голосеменные и занимают господствующее положение на суше. В целом в наземной флоре происходит постепенная смена мезозойской растительности голосеменных (хвойных, цикадовых, гинкговых) растительностью кайнозойского облика. Растительность позднемеловой эпохи уже характеризуется присутствием значительного количества таких современных цветковых растений, как бук, ива, береза, платан, лавр, магнолия. Эта перестройка растительности подготовила хорошую кормовую базу для развития высших наземных позвоночных животных - млекопитающих и птиц. Развитие цветковых растений было связано с расцветом многочисленных насекомых, которые играли важную роль в опылении.
Наступление нового периода в развитии растений не приводило it полному уничтожению древних растительных форм. Часть организмов биосферы сохранялась. Бактерии с появлением цветковых растений не только не исчезли, но продолжали существовать, найдя новые источники питания в почве и в органическом веществе растений и животных. Водоросли разных групп изменялись и развивались наряду с высшими растениями.
Хвойные леса, появившиеся в мезозое, произрастают и сейчас наряду с лиственными. Они дают приют папоротникообразным растениям, так как эти древние обитатели туманного и влажного климата каменноугольного периода боятся открытых мест, освещенных солнцем.
Наконец, следует отметить наличие персистентных форм в составе современной флоры. Наиболее персистентными оказались отдельные группы бактерий, практически не изменившиеся со времени раннего докембрия. Но и из более высокоорганизованных форм растений также образовались роды и виды, которые мало изменились к настоящему времени.

Следует отметить несомненное присутствие в составе современной флоры относительно высокоорганизованных многоклеточных родов растений. Позднепалеозойские и мезозойские формы растений, которые без изменений прожили десятки и сотни миллионов лет, безусловно, относятся к персистентным. Таким образом, в настоящее время среди растительного мира сохранились ''живые ископаемые'' (рис. 15) из групп папоротников, голосеменных и плаунов. Термин ''живое ископаемое'' впервые употребил Ч. Дарвин, указав в качестве примера восточно-азиатское дерево из голосеменных Ginkgo biloba. Из мира наземных растений к живым ископаемым относятся наиболее известные папоротниковые пальмы, гинкговое дерево, араукарии, мамоново дерево, или секвойя.
Как отмечал знаток ископаемой флоры А. Н. Кршптофович, многие роды растений, владыки древних лесов, также существовали чрезвычайно долго, особенно в палеозое; например, Sigillaria, Lepidodendron, Calamites - не менее 100-130 млн лет. Столько жемезозойские папоротники 11 хвойные Metasequoi Род Ginkgo насчитывает более 150 млн лет, а современный вид Ginkgo biloba, если включать в него, по существу, неотличимую форму Ginkgo adiantoides, - около 100 млн лет.
Живые ископаемые современного растительного мира иначе можно назвать филогенетически законсервированными типами. Хорошо изученные в палеоботаническом отношении растения, относимые к живым ископаемым, являются консервативными группами. Они совсем не изменились или изменились очень мало по сравнению с родственными формами геологического прошлого.
Естественно, что наличие живых ископаемых в современной флоре ставит проблему их образования в истории биосферы. Консервативные организации присутствуют во всех крупных филогенетических ветвях и существуют в самых различных условиях: в глубоководных и мелководных зонах моря, в древних тропических лесах, на открытых степных просторах и во всех без исключения водоемах. Важнейшее условие для существования консервативных в эволюционном отношении организмов - наличие мест обитания с постоянной средой жизни. Однако стабильные условия обитания не являются решающими. Присутствие только отдельных форм, а не всех сообществ флоры и фауны указывает на другие факторы сохранности живых ископаемых. Изучение их географического распространения свидетельствует о том, что они приурочены к строго определенным территориям, при этом характерна географическая изоляция. Так, Австралия, острова Мадагаскар и Новая Зеландия - это типичные области распространения наземных живых ископаемых.
В своей эволюции растительный мир создает общий облик древних ландшафтов, в которых происходило развитие животного мира. Поэтому подразделение геологического времени может быть проведено на основании смены различных растительных форм. Немецкий палеоботаник В. Циммерман еще в 1930 г. подразделил все геологическое прошлое с точки зрения развития растительного мира на шесть эр. Он дал им буквенное обозначение и расположил в последовательности от древних эр к более молодым.
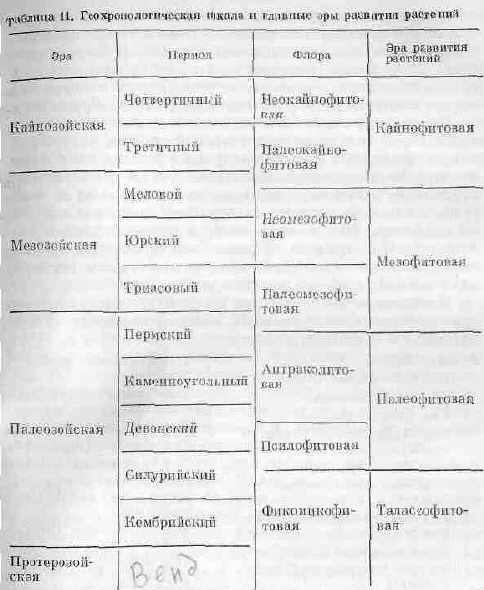
Сопоставление обычной шкалы геологического времени, построенной преимущественна по палеозоологическим данным, со шкалой развития растений представлено в табл. 11.
